Reproductive Toxicity of DDVP: Endocrine Disruption and Gametotoxic Effects
-
Aina Olawale Samson
Department of Physiology, Lagos State University College of Medicine, Ikeja, Lagos State, 23401, Nigeria
Igbayilola Dimeji Yusuff

Department of Human Physiology, College of Medicine and Health Sciences, Baze University, Abuja, Nigeria
| Received 03 Aug, 2025 |
Accepted 10 Oct, 2025 |
Published 31 Dec, 2025 |
A common organophosphate insecticide and pesticide, particularly in poor nations, is Dichlorvos (DDVP). Acetylcholinesterase (AChE) inhibition is its main mode of action, which causes excessive acetylcholine and neurotoxicity. Aside from its neurotoxicity, recent research has shown that DDVP causes serious reproductive damage by disrupting the hormones and showing gametotoxicity. The DDVP alters testicular shape, alters spermatogonial cell viability, decreases testosterone, and alters sperm standard in rodent experiments. Moreover, it inhibits androgen receptor function and may increase susceptibility to prostate cancer in farm workers exposed to it. The DDVP and other Endocrine-Disrupting Chemicals (EDCs) alter hormone signaling in humans and animals repeatedly due to repeated exposure to these chemicals at work, through food, or otherwise. The above disturbances are of concern, especially at the most vulnerable life stages, i.e., pregnancy and puberty. Prenatal DDVP exposure can be transferred from mother to foetus and can influence foetal development, motor function, and neurodevelopment. Recent examination of endocrine disruption mechanisms and gametotoxic effects of DDVP-induced reproductive damage. The well-being of the populace is dependent on understanding and mitigating the impact of DDVP in the process of its unprecedented use and control to prevent extended harm, particularly in the context of timely advancements.
| Copyright © 2025 Samson and Yusuff. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
Dichlorvos (DDVP) is a common organophosphate insecticide that is extensively sold under various trade names and locally known as "Sniper" in Nigeria1. Its quick insecticidal effect makes it suitable for the control of pests in indoor spaces, homes, agricultural farms, veterinary clinics, and even aquaculture units2,3. Despite regulatory restrictions in most nations, DDVP is still utilized in low-resource settings, which is a reason to be concerned about the health and environment4.
Toxicologically, DDVP exerts its toxic effects by inhibiting brain acetylcholinesterase (AChE), causing massive synaptic buildup of acetylcholine. It causes hyperstimulation of the cholinergic pathways and affects both the central and autonomic nervous systems. The presentation includes nausea, sweating, vomiting, muscle weakness, fasciculations, convulsions, and in severe cases, coma or death5.
Toxicity symptoms typically occur after AChE activity has been reduced by at least 20%. Acute toxicity is most commonly caused by inhalation, due to the volatility of DDVP, but chronic exposure or repeated exposure can cause the same neurotoxic effects6.
Besides neurotoxicity, organophosphate pesticides like DDVP may also disrupt endocrine function. The agents have been found to disrupt thyroid hormones-thyroxine (T4) and triiodothyronine (T3)-which are accountable for key roles in metabolism, energy homeostasis, and neurological processes7. Disruption of the hormones leads to hypothyroidism and associated neurological consequences such as depression and lethargy8.
Reproductive health is also a matter of concern. DDVP and other insecticides have been reported to suppress ovarian function, reduce corpora lutea, and also interrupt the estrous cycle in laboratory models9. These are pivotal organs for progesterone secretion, essential for pregnancy and control of menstruation. Some other organophosphates and organochlorines, such as DDT and methamidophos, have been found to possess estrogenic activity that inhibits Gonadotropin-Releasing Hormone (GnRH), finally inhibiting the release of Follicle-Stimulating Hormone (FSH) and Luteinizing Hormone (LH)10. In males, pesticide exposure has been linked with reduced testosterone levels and testicular damage. For instance, 3,5,6-Trichloro-2-Pyridinol (TCPY), a metabolite of chlorpyrifos, and fipronil have both been seen to dampen hormonal balance and spermatogenesis11. In experimental animal models, DDVP has been demonstrated to cause systemic and organ-specific toxicities that impact the inflammatory, metabolic, haematological, hepatic, and cardiovascular systems12-16.
Despite the widespread use of DDVP, its precise mechanisms on reproductive hormones and the viability of germ cells are not well researched. While its neurotoxicity and enzyme inhibition mechanism are reasonably well researched, limited information is available regarding its gametotoxicity and endocrine-disrupting activity in mammals. Therefore, this study aims to determine the reproductive effects of exposure to DDVP based on hormonal regulation, morphology of reproductive organs, and germ cell integrity.
MECHANISM OF ACTION
Acetylcholinesterase (AChE), a neurotransmission enzyme, hydrolyzes acetylcholine in the synaptic cleft17. DDVP inhibits AChE by phosphorylating the active serine residue, creating a stable complex that prevents enzymatic activity17. This inhibition increases synaptic acetylcholine, causing overrelease of glutamate and potentially sustaining status epilepticus18. For example, monocrotophos rapidly inhibits the action of AChE in the hippocampus and striatum. Insecticides such as deltamethrin and benzoylphenyl urea also inhibit AChE activity19.
Allethrin increases SH-SY5Y neuroblastoma cells’ Reactive Oxygen Species (ROS), leading to oxidative stress and cellular damage. Chlorpyrifos (CPF) also significantly compromises genetic and neurological integrity by causing widespread DNA damage and suppressing the activities of AChE, Butyrylcholinesterase (BChE), and carboxylesterase (CbE)20. Moreover, a recent report discovered that DDVP induces necrotic cell death in H9C2 cells by promoting ER stress. However, SIRT1, an inducer of autophagy, safeguards H9C2 cells from the toxicity of DDVP. In the process, it blocks necroptosis via the inhibition of excessive ROS generation and ER stress. Furthermore, low-level exposure to permethrin insecticide at early stages of life causes persistent effects, such as hypotrophy of the heart, along with increased expression of the Nrf2 gene and calcium (Ca2+) in later life21. All things being equal, different pesticides cause cardiac toxicity through the disruption of the heart’s function, which eventually results in death Fig. 1.
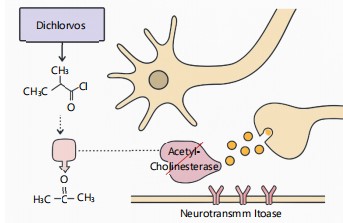
|
Figure 1 illustrates how the organophosphate insecticide dichlorvos (DDVP) interferes with regular neuronal and hormonal processes. Acetylcholine builds up at cholinergic synapses as a result of DDVP’s inhibition of acetylcholinesterase (AChE), which results in ongoing nerve stimulation and damage. By attaching to androgen and oestrogen receptors, DDVP imitates or inhibits endogenous hormones in reproductive organs, interfering with hormonal signalling and gene expression. Developmental and reproductive toxicity may arise from this interference’s effects on spermatogenesis, folliculogenesis, and the hypothalamic-pituitary-gonadal axis. The picture also emphasizes how DDVP can damage DNA and induce oxidative stress, which adds to its genotoxic effects.
DDVP AND ENDOCRINE DISRUPTION
DDVP is widely used in veterinary, agricultural, and domestic settings for pest control6. Despite its established neurotoxicity through inhibition of acetylcholinesterase, recent research shows that DDVP is also an endocrine-disrupting chemical (EDC) that interferes with hormone signaling pathways22. It can imitate, inhibit, or modify the Hypothalamic-Pituitary-Gonadal (HPG) axis, leading in its way to disturbed reproductive hormone patterns, pathological development, and gonadal dysfunction. Experimental studies have revealed that DDVP affects both the estrogenic and androgenic pathways, influencing hormone production, sexual differentiation, and reproductive organ development23.
Mechanistically, DDVP works to interfere with endocrine function by binding hormone receptors and deregulating steroidogenic enzyme expression. This interference lowers circulating concentrations of primary sex hormones such as testosterone and estradiol, which can slow puberty, inhibit fertility, and change reproductive behavior24. Due to its environmental stability and bioaccumulation, chronic low-dose DDVP exposure is a significant reproductive and developmental risk25. It is typical of the overall public health concerns of widespread use of endocrine-disrupting pesticides.
Androgenic activities of DDVP: To enable normal male sexual development, including masculinization of the brain and reproductive tract, fetal testes begin testosterone production around gestational day 6526. Testosterone is central to male physiology, regulating spermatogenesis, testicular and accessory gland development, and maintenance of secondary sexual features like muscle composition and bone mineral content. The male brain also develops under the influence of estradiol, formed by aromatization of testosterone by the Enzyme Aromatase (CYP19). The action of testosterone and its more potent metabolite, Dihydrotestosterone (DHT), is through androgen receptors (ARs), which are expressed in varied tissues including the brain, pituitary, and reproductive organs27.
Perinatal exposure to DDVP may disrupt this finely regulated androgen-dependent process. Interference with testosterone production, AR binding, or subsequent gene activation can disrupt androgen signaling, leading to partial masculinization or feminization of genetically male fetuses28. Disruptions of this nature might have long-term consequences on the Hypothalamic-Pituitary-Gonadal (HPG) axis and impair male reproductive development23. The DDVP may also reduce AR protein levels or interfere with luteinizing hormone stimulation, ultimately affecting feedback loops of testosterone28,29.
Osteogenic activities of DDVP: The 17β-estradiol (E2) plays a critical role in regulating numerous biological processes in both women and men. In women, it is involved in brain, bone, cardiovascular, and reproductive organ development, and menstrual cycle, and pregnancy-induced physiological changes30. In men, E2 is implicated in the development and functioning of reproductive and non-reproductive organs. E2 exerts its effects by binding with ERα and ERβ, ligand-activated transcription factors that possess distinct, tissue-specific activities. ERα is found predominantly in the uterus, breast, testes, and pituitary, while ERβ is more diffusely distributed among tissues. Though co-expressed in the majority of organs, a receptor type will usually dominate and produce a specific biological response31.
Xenoestrogens are endocrine-disrupting chemicals that act by competing with or intercalating with indigenous estrogens by binding to ERs. DES, BPA, and genistein are compounds exhibiting receptor subtype selectivity, which determines tissue-specific activity. Of specific interest, DDVP, while a major organophosphate pesticide, was discovered to be an estrogenic agent by ER-mediated pathways32. DDVP too can mimic E2, alter transcription of genes in estrogen-sensitive tissues, and cause developmental and reproductive toxicity33.
GAMETOTOXICITY OF DDVP
Hypothalamic-pituitary-gonadal axis: In both males and females, the Hypothalamic-Pituitary- Gonadal (HPG) axis is crucial for controlling reproductive function. Steroid hormones like testosterone, oestrogen, and progesterone; regulatory proteins like activins, inhibins, and follistatin; and peptide hormones like Gonadotropin-Releasing Hormone (GnRH), Luteinizing Hormone (LH), and Follicle-Stimulating Hormone (FSH) are all part of this hormonal signalling network34. The hypothalamus secretes GnRH, which then makes its way to the anterior pituitary via the hypophyseal portal system, where it causes the release of FSH and LH. The blood carries LH and FSH to the gonads, where they control the creation of sex steroids and gametogenesis35.
The LH and FSH exhibit sexually dimorphic actions. In males, LH stimulates Leydig cells to release testosterone and FSH stimulates Sertoli cells to assist spermatogenesis. In females, LH stimulates theca cells to release androgens and progesterone production in granulosa cells, while FSH stimulates follicular maturation and estradiol production by aromatase activity36. The process operates on a feedback system where rising levels of testosterone, estrogen, and inhibin regulate the hypothalamus and pituitary to maintain hormonal equilibrium.
The DDVP interferes with these delicate feedback mechanisms by either mimicking or competing with endogenous hormones. DDVP can disrupt androgen and estrogen signaling, influencing gametogenesis, fertility, and hormone regulation37. In men, it has also been linked to reduced testosterone secretion and morphological reproductive alterations. In women, exposure to DDVP results in anovulation, irregular cycles, and accelerated reproductive senescence38. These signs are particularly significant during prenatal or early postnatal development, where exposure to DDVP leads to long-term reproductive and neuroendocrine impairment. The DDVP also has genotoxic and epigenetic effects. DDVP generates Reactive Oxygen Species (ROS), which induce oxidative stress and DNA damage39. Elevated DNA adducts, cell cycle arrest, and chromosomal abnormalities are generated upon DDVP exposure in vitro40. Genotoxicity is yet to be conclusively established in vivo, but these findings demonstrate the capacity of DDVP to induce genetic stability disruption, resulting in reproductive toxicity and disease41 Fig. 2.
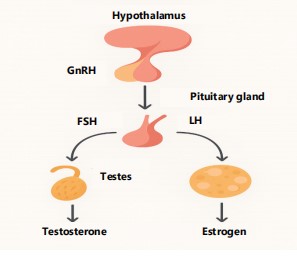
|
The HPG axis and its function in regulating reproduction are depicted in this diagram. Gonadotropin-Releasing Hormone (GnRH), which is secreted by the hypothalamus, causes the anterior pituitary to release Follicle-Stimulating Hormone (FSH) and Luteinizing Hormone (LH). The gonads (ovaries or testes) receive these hormones through the bloodstream, where they control the creation of sex steroids (progesterone, estrogen, and testosterone) and gametogenesis. In order to preserve hormonal equilibrium, these steroids subsequently have an impact on the pituitary and hypothalamus. The diagram further supports the axis’s finely calibrated feedback processes by highlighting the roles that follistatin, activin, and inhibin play in regulating gonadotropin levels.
DEVELOPMENTAL AND TRANSGENERATIONAL EFFECTS
Although many animal studies have been carried out on this topic, the impact of DDVP exposure on human development has not been investigated in any research. McLachlan42 investigated the impact of varying DDVP concentrations on zebrafish embryonic and larval development and documented problems such as heart oedema, loss of blood circulation, delayed hatching, and vertebral malformations. The effects of inhalation of DDVP on the development of 15 pregnant E rats during their 20-gestation period were also investigated42. They observed skeletal defects, gastroschisis, sites of resorption, stillbirths, and other external malformations.
EXPERIMENTAL STUDIES
Humans: The impacts of DDVP on human reproduction are not well established in the literature43.
Experimental investigations: In a previous study, Sherman rats were given an intraperitoneal injection of DDVP at 15 mg/kg body weight in peanut oil on gestation day 11. On gestation day 20, there were no dichotomies in m aternal weight gain, number of fetuses per litter, placental or fetal weight, or number of resorptions per dam44,45. However, the treated group had 41 offspring, three of whom exhibited omphaloceles-a condition that is not present in the control group. Low doses of DDVP, when used with the carcinogen N-methyl-N-nitrosourea, induced ventral prostate alterations in rats in terms of structure and lipid metabolism that may be a causative factor for prostate tumors28. The Hypothalamic-Pituitary-Testicular (HPT) axis was significantly suppressed after exposure, as evidenced by dramatic drops in hormone levels in the blood. In particular, testosterone levels significantly decreased from 6.01±0.50 to 0.74±0.05 (p<0.0001), Luteinizing Hormone (LH) levels decreased from 46.38±1.38 to 19.00±0.46 (p<0.0001), and Follicle-Stimulating Hormone (FSH) levels decreased from 60.00±1.04 to 21.13±0.52 (p<0.0001)46.
Although no visible structural abnormalities were observed in brain morphology on light microscopy at birth, retarded development and immaturity of synaptic connections in the motor cortex have been described using electron microscopy. The effects of prolonged exposure to DDVP vapor were also studied: Carworth E rats and Dutch rabbits were exposed 23 hours a day during gestation to up to 6.25 mg/m³ and 4 mg/m³, respectively. Though these exposures did not affect litter size, gestation rates, resorptions, or foetal weights, they did produce a dose-related reduction in plasma, red blood cell, and brain tissue cholinesterase activity43. DDVP has also been shown to induce subacute and subchronic reproductive toxicity effects47. Conversely, reproductive performance remained unaffected in pregnant sows on DDVP in polyvinyl chloride formulations at 5 or 25 mg/kg bwt during the last 30 days of gestation despite increased fetal brain acetylcholinesterase and decreased maternal plasma, red cell, and myometrial cholinesterase activity at higher doses48. No effects on development or reproduction were noted in orally fed rabbits with 34 mg/kg or pigs fed up to 500 ppm via diet for 37 months.
REGULATORY PERSPECTIVES AND RISK MITIGATION ON DDVP
Dichlorvos (DDVP) has been the focus of significant regulatory scrutiny due to its high acute toxicity and environmental persistence. Classified by the World Health Organization as a Class 1B "highly hazardous" pesticide2, DDVP has been restricted or banned in several countries. The U.S. Environmental Protection Agency (USEPA) cancelled certain residential and agricultural uses of DDVP based on health risk assessments that linked the chemical to neurological and reproductive toxicity, particularly in vulnerable populations such as children and pregnant women4. Similarly, the European Union does not permit DDVP use within its member states, citing unacceptable risks to human health and the environment. However, in many developing countries, DDVP remains readily available and widely used due to weak regulatory enforcement and a lack of safer, affordable alternatives.
Risk mitigation strategies for DDVP often involve promoting safer handling practices, limiting indoor applications, and substituting less toxic alternatives. In regions where DDVP is still permitted, regulatory bodies recommend the use of personal protective equipment (PPE) during application, adherence to recommended dosage, and proper storage away from food or water supplies. Public health campaigns have also played a role in educating communities about the dangers of misuse, particularly in areas where DDVP is repurposed as a suicide agent or indiscriminately used in household pest control. Additionally, international agencies such as the FAO and WHO have guided integrated pest management (IPM) strategies that reduce dependence on chemical pesticides by emphasizing biological control methods and environmental sanitation.
Effective risk mitigation must also involve strengthening regulatory frameworks, including the classification, labeling, and sale of hazardous pesticides. Governments and public health institutions must collaborate to implement monitoring programs that track pesticide residues in food, water, and the environment. Enhancing laboratory capacity and surveillance systems can help identify exposure risks and assess compliance with safety standards. Moreover, investments in research and development of less hazardous pest control methods can facilitate the gradual phase-out of DDVP in favor of safer alternatives. Ultimately, a coordinated global approach that integrates regulation, education, and innovation is essential to mitigate the risks associated with DDVP exposure while ensuring effective pest management in both agricultural and domestic contexts.
CONCLUSION
Pesticides such as DDVP are now being used worldwide on a large scale in various sectors, including healthcare and agriculture. Ever since its extensive use, there is a significant risk of exposure to DDVP by human and animal subjects, which has adverse effects on health. Exposure to DDVP has been linked to several health hazards, including pulmonary, hepatic, cardiac, and neurotoxicity, as well as additional effects on the skin and eyes. Additionally, DDVP has been directly linked to issues related to reproduction, including infertility in men. Researchers have discovered ways that pesticides harm human health by imitating hormones.
Little research is available, however, explaining the molecular mechanisms through which DDVP affects male and female reproduction. Although interference with the PI3K/AKT pathway by some pesticides has been noted to affect sperm, more studies need to elucidate the processes underlying this at a deeper molecular level. There are limitations in the sheer size of the body of the literature available, but this review gives a glimpse of the health issues pertaining to exposure in the context of endocrine disruption and Gametotoxic effects. The implications also highlight the importance of the priority given to health education, research studies, and legislative action to ably counter the negative impact of DDVP on reproductive health. In order to promote their judicious use and become familiar with the health problems of DDVP exposure, it is essential to know these mechanisms.
SIGNIFICANCE STATEMENT
This study discovered the endocrine-disrupting and Gametotoxic Potential of Dichlorvos (DDVP), revealing its capacity to disrupt the hypothalamic-pituitary-gonadal axis, alter reproductive hormone levels, and impair gamete quality through oxidative stress-induced DNA damage. These findings can be beneficial for toxicologists, public health authorities, and policymakers in understanding the mechanisms by which DDVP compromises reproductive health and fertility. The study further highlights the risks of long-term and transgenerational reproductive harm in populations with significant environmental or occupational exposure to organophosphate pesticides. This study will help the researchers to uncover the critical areas of pesticide-induced reproductive toxicity that many researchers were not able to explore. Thus, a new theory on organophosphate-mediated endocrine and genetic disruption may be arrived at.
REFERENCES
- Abaukaka, Y.A., S. Sanusi, K.A. Ozigi and F.U. Malo, 2020. Assessment of the cytotoxic and mutagenic potential of dichlorvos (DDVP) using in silico classification model; a health hazard awareness in Nigeria. Environ. Anal. Health Toxicol., 35.
- Pathak, V.M., V.K. Verma, B.S. Rawat, B. Kaur and N. Babu et al., 2022. Current status of pesticide effects on environment, human health and it’s eco-friendly management as bioremediation: A comprehensive review. Front. Microbiol., 13.
- Binukumar, B.K., A. Bal, R.J.L. Kandimalla and K.D. Gill, 2010. Nigrostriatal neuronal death following chronic dichlorvos exposure: Crosstalk between mitochondrial impairments, α synuclein aggregation, oxidative damage and behavioral changes. Mol. Brain, 3.
- Li, S.S., S.M. Fang, J. Chen, Z. Zhang and Q.Y. Yu, 2023. Effects of short-term exposure to volatile pesticide dichlorvos on the olfactory systems in Spodoptera litura: Calcium homeostasis, synaptic plasticity and apoptosis. Sci. Total Environ., 864.
- Aroniadou-Anderjaska, V., T.H. Figueiredo, M. de Araujo Furtado, V.I. Pidoplichko and M.F.M. Braga, 2023. Mechanisms of organophosphate toxicity and the role of acetylcholinesterase inhibition. Toxics, 11.
- Okoroiwu, H.U. and I.A. Iwara, 2018. Dichlorvos toxicity: A public health perspective. Interdiscip. Toxicol., 11: 129-137.
- Uwamahoro, C., J.H. Jo, S.I. Jang, E.J. Jung, W.J. Lee, J.W. Bae and W.S. Kwon, 2024. Assessing the risks of pesticide exposure: Implications for endocrine disruption and male fertility. Int. J. Mol. Sci., 25.
- Sharma, N., D. Garg, R. Deb and R. Samtani, 2017. Toxicological profile of organochlorines aldrin and dieldrin: An Indian perspective. Rev. Environ. Health, 32: 361-372.
- Pascotto, V.M., M.T. Guerra, J.A.A. Franci, J.L.V. de Camargo, W.G. Kempinas and C.A.S. Franchi, 2015. Effects of a mixture of pesticides on the adult female reproductive system of Sprague-Dawley, Wistar, and Lewis rats. J. Toxicol. Environ. Health Part A, 78: 602-616.
- Yoon, K., S.J. Kwack, H.S. Kim and B.M. Lee, 2014. Estrogenic endocrine-disrupting chemicals: Molecular mechanisms of actions on putative human diseases. J. Toxicol. Environ. Health Part B, 17: 127-174.
- de Barros, A.L., J.H. Bae, C.S. Borges, J.L. Rosa and M.M. Cavariani et al., 2016. Perinatal exposure to insecticide fipronil: Effects on the reproductive system in male rats. Reprod. Fertil. Dev., 29: 1130-1143.
- Saka, W.A., Y.D. Igbayilola, H.J. Lawan, M.B. Zakari, D.E. Awujoola, P.O. Olarinde and V.O. Adegoke, 2025. L-arginine supplement ameliorates dichlorvos-induced systemic inflammatory response and liver dysfunction in male Wistar rats. Toxicol. Rep., 14.
- Saka, W.A., F.M. Moronkeji, F.M. Akano, Y.D. Igbayilola, L.J. Hamidu and M.B. Zakari, 2025. Methanol leaf extract of Ricinus communis L. mitigates dichlorvos-induced cardiac injury via anti-inflammatory, antioxidant, and anti-apoptotic mechanisms in Wistar rats. Trop. J. Nat. Prod. Res., 9: 2271-2278.
- Adeoye, S.W., O. Seun, I.Y. Dimeji, O.E. Ugochukwu and H.L. Jabba, 2025. Hepatoprotective effects of curcumin against dichlorvos-induced toxicity in male Wistar rats. J. Appl. Life Sci. Int., 28: 41-54.
- Adeoye, S.W., I.Y. Dimeji, H.L. Jabba, N. Murtala and Z.M. Baba et al., 2025. Curcumin mitigates dichlorvos-induced organ toxicity, restoring hematological and metabolic functions. Pharmacol. Res. Nat. Prod., 8.
- Saka, W.A., Y.D. Igbayilola, J.H. Lawan, T.K. Muftaudeen and R. Adejumo et al., 2024. L-arginine supplementation mitigates dichlorvos-induced haematocardiotoxicity, and oxidative stress in male Wistar rats. Open J. Appl. Sci., 14: 1886-1903.
- Aroniadou-Anderjaska, V., J.P. Apland, T.H. Figueiredo, M. de Araujo Furtado and M.F. Braga, 2020. Acetylcholinesterase inhibitors (nerve agents) as weapons of mass destruction: History, mechanisms of action, and medical countermeasures. Neuropharmacology, 181.
- Figueiredo, T.H., J.P. Apland, M.F.M. Braga and A.M. Marini, 2018. Acute and long-term consequences of exposure to organophosphate nerve agents in humans. Epilepsia, 59: 92-99.
- Chen, J., X. Ye, J. Wang, B. Xia and T. Xin, 2020. Transcriptome analysis of Tetranychus cinnabarinus responses to an insecticide exposure. Syst. Appl. Acarol., 25: 1329-1342.
- Muller, M., L. Hess, A. Tardivo, R. Lajmanovich and A. Attademo et al., 2014. Neurologic dysfunction and genotoxicity induced by low levels of chlorpyrifos. NeuroToxicology, 45: 22-30.
- Vadhana, M.S.D., S.S. Arumugam, M. Carloni, C. Nasuti and R. Gabbianelli, 2013. Early life permethrin treatment leads to long-term cardiotoxicity. Chemosphere, 93: 1029-1034.
- Tao, Y., Z. Li, Y. Yang, Y. Jiao, J. Qu, Y. Wang and Y. Zhang, 2022. Effects of common environmental endocrine-disrupting chemicals on zebrafish behavior. Water Res., 208.
- Pan, J., P. Liu, X. Yu, Z. Zhang and J. Liu, 2024. The adverse role of endocrine disrupting chemicals in the reproductive system. Front. Endocrinol., 14.
- Srnovršnik, T., I. Virant-Klun and B. Pinter, 2023. Polycystic ovary syndrome and endocrine disruptors (bisphenols, parabens, and triclosan)-A systematic review. Life, 13.
- Shekhar, C., R. Khosya, K. Thakur, D. Mahajan, R. Kumar, S. Kumar and A.K. Sharma, 2024. A systematic review of pesticide exposure, associated risks, and long-term human health impacts. Toxicol. Rep., 13.
- Nef, S. and L.F. Parada, 2000. Hormones in male sexual development. Genes Dev., 14: 3075-3086.
- Alemany, M., 2022. The roles of androgens in humans: Biology, metabolic regulation and health. Int. J. Mol. Sci., 23.
- Quintino-Ottonicar, G.G., L.R. da Silva, V.L.R. da Silva Maria, E.M. Pizzo and A.C.P. de Santana et al., 2023. Exposure to Dichlorvos pesticide alters the morphology of and lipid metabolism in the ventral prostate of rats. Front. Toxicol., 5.
- Krause, W., 1977. Influence of DDT, DDVP and malathion on FSH, LH and testosterone serum levels and testosterone concentration in testis. Bull. Environ. Contam. Toxicol., 18: 231-242.
- Bartkowiak-Wieczorek, J., A. Jaros, A. Gajdzińska, P. Wojtyła-Buciora and I. Szymański et al., 2024. The dual faces of oestrogen: The impact of exogenous oestrogen on the physiological and pathophysiological functions of tissues and organs. Int. J. Mol. Sci., 25.
- Chen, P., B. Li and L. Ou-Yang, 2022. Role of estrogen receptors in health and disease. Front. Endocrinol., 13.
- Vandenberg, L.N., A. Najmi and J.P. Mogus, 2020. Agrochemicals with estrogenic endocrine disrupting properties: Lessons learned? Mol. Cell. Endocrinol., 518.
- Dutta, S., P. Sengupta, S. Bagchi, B.S. Chhikara, A. Pavlík, P. Sláma and S. Roychoudhury, 2023. Reproductive toxicity of combined effects of endocrine disruptors on human reproduction. Front. Cell Dev. Biol., 11.
- Acevedo‐Rodriguez, A., A.S. Kauffman, B.D. Cherrington, C.S. Borges, T.A. Roepke and M. Laconi, 2018. Emerging insights into hypothalamic-pituitary-gonadal axis regulation and interaction with stress signalling. J. Neuroendocrinol., 30.
- Casati, L., S. Ciceri, R. Maggi and D. Bottai, 2023. Physiological and pharmacological overview of the gonadotropin releasing hormone. Biochem. Pharmacol., 212.
- Casarini, L., D. Santi, G. Brigante and M. Simoni, 2018. Two hormones for one receptor: Evolution, biochemistry, actions, and pathophysiology of LH and hCG. Endocr. Rev., 39: 549-592.
- de Coster, S. and N. van Larebeke, 2012. Endocrine-disrupting chemicals: Associated disorders and mechanisms of action. J. Environ. Public Health, 2012. .
- Rattan, S., C. Zhou, C. Chiang, S. Mahalingam, E. Brehm and J.A. Flaws, 2017. Exposure to endocrine disruptors during adulthood: Consequences for female fertility. J. Endocrinol., 233: R109-R129.
- Intidhar Ben Salem, M. Boussabbeh, I. Graiet, A. Rhouma, H. Bacha and S.A. Essefi, 2016. Quercetin protects HCT116 cells from dichlorvos-induced oxidative stress and apoptosis. Cell Stress Chaperones, 21: 179-186.
- Fiore, M., M. Mattiuzzo, G. Mancuso, P. Totta and F. Degrassi, 2013. The pesticide dichlorvos disrupts mitotic division by delocalizing the kinesin Kif2a from centrosomes. Environ. Mol. Mutagen., 54: 250-260.
- Trivedi, S.P., A. Ratn, Y. Awasthi, M. Kumar and A. Trivedi, 2021. In vivo assessment of dichlorvos induced histological and biochemical impairments coupled with expression of p53 responsive apoptotic genes in the liver and kidney of fish, Channa punctatus (Bloch, 1793). Comp. Biochem. Physiol. C: Toxicol. Pharmacol., 245.
- McLachlan, J.A., 2006. Commentary: Prenatal exposure to diethylstilbestrol (DES): A continuing story. Int. J. Epidemiol., 35: 868-870.
- Thorpe, E., A.B. Wilson, K.M. Dix and D. Blair, 1972. Teratological studies with dichlorvos vapour in rabbits and rats. Arch. Toxikologie, 30: 29-38.
- Lazarini, C.A., R.Y. Lima, A.P. Guedes and M.M. Bernardi, 2004. Prenatal exposure to dichlorvos: Physical and behavioral effects on rat offspring. Neurotoxicol. Teratol., 26: 607-614.
- Imam, A., N.A. Sulaiman, A.L. Oyewole, S. Chengetanai and V. Williams et al., 2018. Chlorpyrifos- and dichlorvos-induced oxidative and neurogenic damage elicits neuro-cognitive deficits and increases anxiety-like behavior in wild-type rats. Toxics, 6.
- Saka, W.A., A.A. Oladipo, O.R. Kolawole, A. Olayioye and R.E. Akhigbe, 2025. Sexual dysfunction in dichlorvos-exposed male Wistar rat is ameliorated by curcumin and associated with the upregulation of testosterone. Naunyn-Schmiedeberg's Arch. Pharmacol., 398: 1003-1012.
- Dirican, E.K. and Y. Kalender, 2012. Dichlorvos-induced testicular toxicity in male rats and the protective role of vitamins C and E. Exp. Toxicol. Pathol., 64: 821-830.
- Stanton, H.C., J.R. Albert and H.J. Mersmann, 1979. Studies on the pharmacology and safety of dichlorvos in pigs and pregnant sows. Am. J. Vet. Res., 40: 315-320.
How to Cite this paper?
APA-7 Style
Samson,
A.O., Yusuff,
I.D. (2025). Reproductive Toxicity of DDVP: Endocrine Disruption and Gametotoxic Effects. Trends in Pharmacology and Toxicology, 1(2), 103-111. https://doi.org/10.21124/tpt.2025.103.111
ACS Style
Samson,
A.O.; Yusuff,
I.D. Reproductive Toxicity of DDVP: Endocrine Disruption and Gametotoxic Effects. Trends Pharm. Toxicol. 2025, 1, 103-111. https://doi.org/10.21124/tpt.2025.103.111
AMA Style
Samson
AO, Yusuff
ID. Reproductive Toxicity of DDVP: Endocrine Disruption and Gametotoxic Effects. Trends in Pharmacology and Toxicology. 2025; 1(2): 103-111. https://doi.org/10.21124/tpt.2025.103.111
Chicago/Turabian Style
Samson, Aina, Olawale, and Igbayilola Dimeji Yusuff.
2025. "Reproductive Toxicity of DDVP: Endocrine Disruption and Gametotoxic Effects" Trends in Pharmacology and Toxicology 1, no. 2: 103-111. https://doi.org/10.21124/tpt.2025.103.111

This work is licensed under a Creative Commons Attribution 4.0 International License.